Хламидии микробиология характеристика
1. Общая характеристика хламидий
Хламидии – мелкие грамотрицательные бактерии, которые являются облигатными внутриклеточными паразитами, подобно вирусам, но отличаются от них тем, что имеют в своем составе и РНК и ДНК, рибосомы, клеточную стенку и размножаются бинарным делением. Однако, они отличаются от большинства типичных бактерий тем, что они не имеют пептидогликана в клеточной стенке и неспособны образовывать собственную АТФ. Они требуют использования АТФ хозяина – следовательно, являются энергетическими паразитами. Род Chlamydia - единственный в семействе Chlamydiaceae и порядке Chlamydiales.
Существуют три вида, утвержденных международным комитетом: Chlamydia trachomatis. C. psittaci и C. pneumoniae.
C. trachomatis может быть разделена на три биовара:
1. Вызывающий трахому и конъюнктивит с включениями.
2. Вызывающий венерическую лимфогранулему (паховый лимфогрануломатоз).
3. Вызывающий пневмонию у мышей.
Оба биовара, вызывающие заболевания у человека, могут быть подразделены на 17 сероваров. Классическая трахома вызывается сероварами A, Ba, B и C, в то время как конъюнктивит с включениями и генитальныеинфекции - серовары D-K. Венерическую гранулему вызывают, по крайней мере, три серовара (L1, L2, L3).
C. psittaci инфицирует млекопитающих и птиц и хотя имеются и серологические различия и различия в нуклеиновых кислотах, этот вид пока еще не был подразделен. C. pneumomae,, самый новый вид, во-видимому, инфицирует только людей и пока проявляет себя серологически однородным.
Морфология. Хламидии – мелкие неподвижные бактерии, размерами 300-350 нм, которые плохо окрашиваются по Граму. Однако, они хорошо выявляются в препаратах, окрашенных по Романовскому-Гимзе или другими методами в виде частиц, представляющих внеклеточную, инфекционную, метаболически инертную форму хламидий, известную как элементарное тельце. Как только микроорганизм инфицирует клетку хозяина, он увеличивается в размере до 800-1000 нм. Эта неинфекционная но метаболически активная частица известна как ретикулярное тельце.
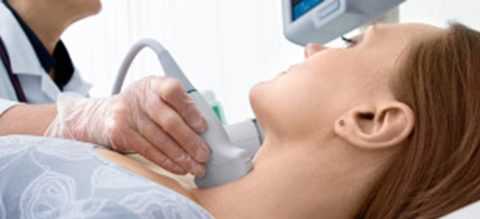
Хламидии могут также обнаруживаться иммунофлюоресцентным методом, когда специфическое антитело, меченое изотиоцианатом флюоресцеина наносится на препараты, приготовленные из клинического материала, например из шейки матки, уретры или из инфицированной культуры клеток. Элементарные тельца выглядят как отдельные ярко-зеленые частицы. С помощью этой методики можно подтвердить хламидийную инфекцию в течение 30 минут после получения материала.
Три вида хламидий можно дифференцировать по:
1. Росту;
2. Профилю нуклеиновой кислоты;
3. Наличию плазмид;
4. Экспрессированным антигенам
5. Включениям в клетки хозяина, которые различаются морфологически.
C. trachomatis образует в своих включениях гликогеновый матрикс, который окрашивается йодом в коричневый цвет, что имеет значение в диагностике.
Элементарные тельца C. trachomatis и C. psittaci 300-350 нм в диаметре с очень небольшим периплазматическим пространством. C. pneumoniae отличается тем, что имеет большое периплазматическое пространств, давая элементарные тела грушевидной формы (310 x 400 нм).
Ретикулярные тельца всех трех видов хламидий крупные, округлой формы (800-1000 нм). Поскольку ретикулярные тельца метаболически активны, их внутренняя структура отражает стадию размножения. В начале цикла обнаруживается много рибосом. Позже ДНК организуется в ядра для потомства элементарных телец. Образование гликогена в ретикулярных тельцах C. trachomatis обнаруживается в электронном микроскопе примерно через 18 часов после заражения, и включения заметны примерно через 22 ч. В случае C. psittaci или C. pneumoniae образования гликогена не отмечается.
Между цитоплазматической мембраной и клеточной стенкой не отмечается наличия наличия пептидогликана, как это характерно для грамотрицательных бактерий.
Нуклеиновая кислота. Молекулярная масса нуклеиновой кислоты хламидий составляет порядка 660 x 106. Три вида хламидий могут быть дифференцированына основании гомологии, поскольку внутривидовое соответствие не превышает 10 %. Плазмидная ДНК обнаружена у большинства штаммов C. trachomatis и C. psittaci, но овечьи штаммы C. psittaci и C. pneumomae, по-видимому, не имеют плазмид.
Белок. Полагают, что небольшое число белков имеет значение для понимания антигенности и патогенеза, а также для возможного получения вкацин.
У хламидий имеется четыре белка наружной мембраны, главный из которых имеет видоспецифические эпитопы. У хламидий имеется также белок (heat shock protein), антитела к которому перекрестно реагируют с подобными белками других микроорганизмов, что делает затруднительной интерпретацию серологического ответа при хламидийных инфекциях.
Липополисахариды (ЛПС). Родо-специфический антиген хламидий может быть получен методами, используемыми для извлечения липополисахаридов из грамотрицательных бактерий. Он имеет три эпитопных участка, из которых только один специфичен для хламидий. Другие эпитопы перекрестно реагируют с мутантами Escherichia coli, Proteus mirabdis и Salmonella typhimunum и с ЛПС, полученным из разных видов Acinetobacter. Поскольку ЛПС-антиген используется в реакции связывания комплемента и некоторых иммуноферментных тестах (ИФА), необходима осторожность при интерпретации результатов.
Культивирование. В 1957 C. trachomatis была выделена путем культивирования в желточном мешке куриного эмбриона. Хотя этот метод теперь не используется широко для выделения хламидий, он применяется для производства антигена, особенно для реакции иммунофлюоресценции. Выделение в культуре клеток было успешно проведено в 1965 г. Для роста хламидий культуры клеток должны быть, чтобы обеспечить продуктивное взаимодействие микроорганизма и клеток-хозяев. Это достигается осаждением микроорганизмов на клетки путем центрифугирования и подавления клеток облучением или добавлением к питательной среде 5-йодо-2’-дезоксиуридина. Окраска по Романовскому-Гимзе выявляет виды хламидий, а для обнаружения содержащих гликоген включений C. trachomatis может быть использован йод. Реакция иммунофлюоресценции с соответствующими антисыворотками позволяет выявить родо- или видо-специфические антигены. Различные виды хламидий имеют незначительно различающиеся требования к условиям культивирования, но C. pneumoniae наиболее трудно культивировать и в куриных эмбрионах, и в культуре клеток.
Цикл развития. Первоначально микроорганизм должен присоединиться к клетке хозяина. Обнаружены специфические прикрепительные белки, что позволяет обсуждать вопрос о возможной роли белковых отростков на поверхности элементарных телец. Не все виды хламидий легко взаимодействуют с клетками хозяина. C. psittaci и хламидии венерической лимфогранулемы присоединяются и покрывают поверхность клетки хозяина, но C. trachomatis и C. pneumoniae вряд ли на это способны. Это зависит частично от отрицательного заряда микроорганизморв и клеток хозяина, котрый приводит к отталкиванию их друг от друга. Для преодоления этого добавляют поликатионы типа ДЭАЭ-декстрана, что увеличивает адсорбцию C. trachomatis, в то время как центрифугирование C. trachomatis или C. pneumoniae с клетками хозяина дает возможность формирования гидрофильных связей и повышает прикрепляемость обоих видов хламидий. Эти взаимодействия, хотя и являются полезными in vitro, вероятно не отражают ситуацию in vivo.
Как только хламидии и клетка хозяина вошли в контакт, микроорганизм поступает в клетку внутри вакуоли. Жизнеспособные хламидии, поступившие в цитоплазму клетки хозяина, подавляют слияние с лизососмами, что позволяет избежать их гибели.
В вакуоле элементарное тельце теряет свое компактное дезоксирибонуклеиновое ядро. Клеточная стенка становится менее твердой из-за нарушения дисульфидных связей; частица увеличивается в размере и становится ретикулярным тельцем. Ретикулярное тельце не имеет цитохрома и неспособно вырабатывать АТФ, снабжение которой должно обеспечиваться хозяином. В вакуоле ретикулярное тельце размножается бинарным делением, в результате чего образуются плеоморфные микроорганизмы. Через 18-20 ч ДНК уплотняется, формируются дисульфидные связи в белках наружной оболочки, и образуются новые элементарные тельца внутри эндосомальных пузырьков Выход элементарных телец идет вследствие лизиса клетки хозяина.
Потребность хламидий в определенных аминокислотах может вести к подавлению размножения, которое может быть восстановлено добавлением необходимых аминокислот.
Иммунитет и патогенез. Хламидийные инфекции вызывают гуморальные и клеточные иммунные реакции, которые играют различную роль в выздоровлении, резистентности к повторному инфицированию и иммунопатологических осложнениях. Хламидийные инфекции у человека не могут в достаточной мере быть изучены экспериментально, так как хотя и были созданы модели на животных, реакции у них могут не вполне соответствовать реакциям у человека. Однако, были использованы две самопроизвольные инфекции, коньюнктивит с включениями у морских свинок и пневмония у мышей. Выздоровление морских свинок замедлялось при подавлении иммунитета в результате введения циклофосфамида или антилимфоцитарной сыворотки. Это указывает на то, что и гуморальный, и клеточный иммунный ответ необходимы для выздоровления от коньюнктивита с включениями у морских свинок.
При инфекциях у человека хламидии обладают способностью к десквамации клеток эпителия, а также макрофагов легких и желудочно-кишечного тракта. Такие инфекции вызывают развитие воспалительной реакции с инфильтрацией нейтрофилами в острой стадии. Внутриклеточное разрушение хламидий нейтрофилами встречается в клетках кислород-зависимыми и кислород-независимыми ферментными системами. Впоследствии в инфильтрате появляются моноциты и лимфоциты. Хламидии не размножаются в моноцитах, но когда эти клетки становятся тканевыми макрофагами, и C. psittaci, и хламидии венерической лимфогранулемы могут высвобождаться из фаголизосом и давать большое количество микроорганизмов. C. trachomatis может также размножаться в макрофагах, но в меньшем объеме. Предполагают, что инфицированные макрофаги могут быть средством распространения хламидий в организме хозяина.
Продукция антител идет в обычной последовательности, вначале IgM, затем - IgG, уровень антител зависит от дозы. Образуется сывороточный IgA, в слизи обнаруживается секреторный IgA (S-IgA). Чем выше уровень S-IgA, тем менее успешно выделение хламидий из полового тракта. Более позднее, на 6-8 неделе, повышение уровня IgM и IgG антител к C. pneumoniae в сравнении с ранним, на 2-3 неделе повышением при генитальной инфекции, необходимо учитывать при серологической диагностике.
Значение секреторных антител (или скорее их недостаточность) хорошо видно у новорожденных, имеющих материнские IgG. У части, инфицированных в родовых путях C. trachomatis, развивается конъюнктивит, несмотря на материнские IgG. Если инфекция не лечится соответствующим образом, у них развивается пневмония в возрасте 4-12 недель, когда уровень материнских антител снижается, и новорожденные начинают образовывать собственные IgM и IgG. Присутствие IgM антител или повышение титров IgG является признаком хламидийной инфекции дыхательного тракта новорожденных.
Хотя специфические антитела могут нейтрализовать хламидии in vitro, существует лишь небольшая защита против повторной инфекции глаз или слизистых половых органов у лиц с циркулирующими IgG антителами. При вторичных или более поздних инфекциях наблюдается очень быстрая инфильтрация нейтрофилами и лимфоцитами. Плазматические клетки образуют антитела, которые не способны привести к выздоровлению. Эта недостаточность защиты антителами имеет значение для попыток разработки эффективной вакцины.
Клеточный ответ. Антитела - не единственный фактор, который нужно рассматривать при повторной инфекции. Клеточно-опосредованный иммунитет также играет роль, которая может быть но может и не быть полезной для организма хозяина.
Цельные живые хламидии, убитые высокой температурой микроорганизмы или даже липополисахарид может стимулировать продукцию лимфокинов типа При трахоме острая инфекция заканчивается воспалительной реакцией с инфильтрацией нейтрофилами и лимфоцитами. Эти лимфоциты могут группироваться и образовывать фолликул. Центр фолликула состоит из B клеток и макрофагов, окруженных T клетками, и все они окружены тонким слоем эпителиальных клеток. Эти эпителиальные клетки слущиваются, поскольку фолликул некротизируется, и регенерация фиброзной ткани, стимулируемая интерлейкином-1, ведет к рубцеванию. При повторной инфекции или повторной стимуляцией хламидийным антигеном воспалительная реакция будет более быстрой, приводящей к формированию большого количества фолликулов и рубцеванию.
То же относится и к половому тракту, где восходящая инфекция может приводить к сальпингиту. Это заканчивается воспалительной реакцией с инфильтрацией нейтрофилами и лимфоцитами, нарушением мерцательного эпителия и эпителиальных клеток вблизи лимфоцитов. Повторяющаяся инфекция вызывает тяжелое поражение труб с формированием фолликулов, значительное разрушение эпителия и рубцевание, ведущее к нарушению проходимости (обструкции) бахромчатого конца труб. Такое поражение может заканчиваться бесплодием из-за нарушения продвижения яйцеклетки и к хронического воспаления тазовых органов.
95. Возбудитель хламидиозов. Таксономия. Характеристика. Микробиологическая диагностика. Лечение.
Таксономия: порядок Chlamydiales, семейство Chlamydaceae, род Chlamydia. Род представлен видами С.trachomatis, C.psittaci,C.pneumoniae.
Болезни, вызываемые хламидиями, называют хламидиозами. Заболевания, вызываемые C. trachomatis u C. pneumoniae, — антропонозы. Орнитоз, возбудителем которого является С. psittaci, — зооантропонозная инфекция.
Морфология хламидий: мелкие, грам «-» бактерии, шаровидной формы. Не образуют спор, нет жгутиков и капсулы. Клеточная стенка: 2-х слойная мембрана. Имеют гликолипиды. По Граму – красный цвет. Основной метод окраски – по Романовскому – Гимзе.
2 формы существования: элементарные тельца (неактивные инфекционные частицы, вне клетки); ретикулярные тельца (внутри клеток, вегетативная форма).
Культивирование: Можно размножать только в живых клетках. В желточном мешке развивающихся куриных эмбрионов, организме чувствительных животных и в культуре клеток
Ферментативная активность: небольшая. Ферментируют пировиноградную кислоту, синтезируют липиды. Не способны синтезировать высокоэнергетические соединения.
Антигенная структура: Антигены трех типов: родоспецифический термостабильный липополисахарид (в клеточной стенке). Выявляют с помощью РСК; видоспецифический антиген белковой природы (в наружной мембране). Обнаруживают с помощью РИФ; вариантоспецифический антиген белковой природы.
Факторы патогенности. С белками наружной мембраны хламидий связаны их адгезивные свойства. Эти адгезины обнаруживают только у элементарных телец. Хламидии образуют эндотоксин. У некоторых хламидий обнаружен белок теплового шока, способный вызывать аутоиммунные реакции.
Резистентность. Высокая к различным факторам внешней среды. Устойчивы к низким температурам, высушиванию. Чувствительны к нагреванию.
С. trachomatis - возбудитель заболеваний мочеполовой системы, глаз и респираторного тракта человека.
Трахома — хроническое инфекционное заболевание, характеризующееся поражением конъюнктивы и роговицы глаз. Антропоноз. Передается контактно- бытовым путем.
Патогенез: поражает слизистую оболочку глаз. Он проникает в эпителий конъюнктивы и роговицы, где размножается, разрушая клетки. Развивается фолликулярный кератоконъюнктивит.
Диагностика: исследование соскоба с конъюнктивы. В пораженных клетках при окраске по Романовскому—Гимзе обнаруживают цитоплазматические включения фиолетового цвета, расположенные около ядра — тельца Провачека. Для выявления специфического хламидийного антигена в пораженных клетках применяют также РИФ и ИФА. Иногда прибегают к культивированию хламидий трахомы на куриных эмбрионах или культуре клеток.
Лечение: антибиотики (тетрациклин) и иммуностимуляторы (интерферон).
Профилактика: Неспецифическая.
Урогенитальный хламидиоз — заболевание, передающееся половым путем. Это — острое/хроническое инфекционное заболевание, которое характеризуется преимущественным поражением мочеполового тракта.
Заражение человека происходит через слизистые оболочки половых путей. Основной механизм заражения — контактный, путь передачи — половой.
Иммунитет: клеточный, с сыворотке инфицированных – специфические антитела. После перенесенного заболевания - не формируется.
Диагностика: При заболеваниях глаз применяют бактериоскопический метод — в соскобах с эпителия конъюнктивы выявляют внутриклеточные включения. Для выявления антигена хламидии в пораженных клетках применяют РИФ. При поражении мочеполового тракта может быть применен биологический метод, основанный на заражении исследуемым материалом (соскобы эпителия из уретры, влагалища) культуры клеток.
Постановка РИФ, ИФА позволяют обнаружить антигены хламидии в исследуемом материале. Серологический метод - для обнаружения IgM против С. trachomatis при диагностике пневмонии новорожденных.
Лечение. антибиотики (азитромицин из группы макролидов), иммуномодуляторы, эубиотики.
Профилактика. Только неспецифическая (лечение больных), соблюдение личной гигиены.
Венерическая лимфогранулема — заболевание, передающееся половым путем, характеризуется поражением половых органов и регионарных лимфоузлов. Механизм заражения — контактный, путь передачи — половой.
Иммунитет: стойкий, клеточный и гуморальный иммунитет.
Диагностика: Материал для исследования - гной, биоптат из пораженных лимфоузлов, сыворотка крови. Бактериоскопический метод, биологический (культивирование в желточном мешке куриного эмбриона), серологический (РСК с парными сыворотками положительна) и аллергологический (внутрикожная проба с аллергеном хламидии) методы.
Лечение. Антибиотики — макролиды и тетрациклины.
Профилактика: Неспецифическая.
С. pneumoniae - возбудитель респираторного хламидиоза, вызывает острые и хронические бронхиты и пневмонии. Антропоноз. Заражение – воздушно-капельным путем. Попадают в легкие через верхние дыхательные пути. Вызывают воспаление.
Диагностика: постановка РСК для обнаружения специфических антител (серологический метод). При первичном заражении учитывают обнаружение IgM. Применяют также РИФ для обнаружения хламидийного антигена и ПЦР.
Лечение: Проводят с помощью антибиотиков (тетрациклины и макролиды).
Профилактика: Неспецифическая.
С. psittaci - возбудитель орнитоза — острого инфекционного заболевания, которое характеризуется поражением легких, нервной системы и паренхиматозных органов (печени, селезенки) и интоксикацией.
Зооантропоноз. Источники инфекции – птицы. Механизм заражения – аэрогенный, путь передачи – воздушно- капельный. Возбудитель – через слиз. оболочки дыхат. путей, в эпителий бронхов, альвеол, размножается, воспаление.
Диагностика: Материал для исследования - кровь, мокрота больного, сыворотка крови для серологического исследования.
Применяют биологический метод — культивирование хламидий в желточном мешке куриного эмбриона, в культуре клеток. Серологический метод. Применяют РСК, РПГА, ИФА, используя парные сыворотки крови больного. Внутрикожная аллергическая проба с орнитином.
Лечение: антибиотики (тетрациклины, макролиды).
16. Хламидии, морфология и другие биологические свойства. Роль в патологии.
Род Chlamydia и род Chlamydophila.
Представители семейства Chlamydiaceae (хламидии) являются патогенными облигатными внутриклеточными бактериями, паразитирующими в чувствительных клетках теплокровных (млекопитающих, птиц, человека и др.).
Они близки по структуре и химическому составу к классическим бактериям. Для них характерно сохранение морфологической сущности на протяжении всего жизненного цикла, деление вегетативных форм, наличие клеточной стенки, содержание ДНК и РНК, энзиматическая активность, чувствительность к антибиотикам широкого спектра, наличие общего родоспецифического антигена.
В то же время хламидии по размерам меньше классических бактерий, имеют небольшой геном, являются облигатными внутриклеточными паразитами с уникальным циклом развития. Они не способны синтезировать высокоэнергетические соединения и обеспечивать собственные энергетические потребности (энергозависимые паразиты), что и определяет их облигатный паразитизм.
С учетом своих особенностей хламидии занимают самостоятельное (особое) положение среди других микроорганизмов — прокариот. Для человека имеют значение преимущественно представители двух родов — Chlamydia и Chlamydophila. Наибольшее значение имеют три вида.
Chlamydia trachomatis , различные серотипы которого вызывают трахому, венерическую лимфогранулему и наиболее распространенные урогенитальные хламидиозы.
Chlamydophila psittaci вызывает орнитоз и зоонозные хламидиозы.
Chlamydophila pneumoniae вызывает антропонозные пневмонии, ОРЗ, с этим возбудителем связывают развитие некоторых форм бронхиальной астмы, атеросклероза.
Морфологические особенности.
Клеточный цикл развития хламидий имеет две основных формы — элементарные тельца (ЭТ) — инфекционная форма и ретикулярные тельца (РТ) — вегетативная форма. Сферические ЭТ значительно меньше размерами (менее 300 нм в диаметре), имеют более жестную электронно- плотную структуру, метаболически мало активны, адаптированы к кратковременному внеклеточному существованию.
Цикл развития хламидий осущесвляется в цитоплазматическом включении — фагосоме (вакуоле), куда ЭТ попадают путем стимулирования эндоцитоза. В процессе адсорбции и эндоцитоза участвуют термолабильные эффекторные белковые поверхностные антигены хламидий. ЭТ подавляют фагосомо — лизосомальное слияние и преобразуются при участии главного поверхностного протеина в РТ, которые обладают активным метаболизмом, более крупными размерами и активным бинарным делением. Цикл размножения заканчивается обратным переходом РТ в ЭТ, разрывом мембран включения и ограничивающих мембран клетки хозяина, выходом ЭТ из клеток, далее ЭТ инфицируют новые клетки. Тельца включений выявляются в клетках при помощи световой и иммунолюминесцентной микроскопии.
Кроме того, хламидии способны образовывать L — формы, персистентные формы.
Культуральные свойства.
Хламидии не растут на питательных средах самого сложного состава (это сближает их по свойствам с риккетсиями и особенно — с вирусами), для их культивирования могут быть использованы лабораторные животные и куриные эмбрионы (биопроба) и особенно чувствительные линии клеток животных — чаще клетки McCoy (чаще с обработкой цитостатиками для повышения чувствительности), которые считались “золотым стандартом” диагностики.
Биохимические свойства.
Способны самостоятельно синтезировать нуклеиновые кислоты, некоторые белки и липиды. Являются энергетически зависимыми от хозяина эндопаразитами. Chlamydia trachomatis обладает гликоген — синтетазной системой (гликоген содержится и выявляется при определенных методах окраски во включениях), синтезирует предшественники фолиевой кислоты и соответственно биохимической активности чувствителен к сульфаниламидам.
Антигенная структура.
1. Выделяют поверхностный родоспецифический антиген (ЛПС), локализующийся на наружной мембране клеточной стенки. ЛПС имееи две антигенные детерминанты, одна из них специфична для всего рода, другая перекрестно реагирует с некоторыми другими грамотрицательными бактериями (сальмонеллы серовара minnesota), термостабилен.
2. МОМР — главный белок наружной мембраны несет функцию структурного белка и порина. Включает термолабильные белковые детерминанты , обладающие видо — , типо — и сероварной специфичностью.
Детерминанты патогенности.
1. Эндотоксин (липополисахарид), подобный эндотоксинам грамотрицательных бактерий.
2. Экзотоксины — термолабильные субстанции.
3. Антигены клеточной поверхности, подавляющие защитные реакции.
Факторы патогенности хламидий препятствуют фагосомо — лизосомальному слиянию в фагоцитах.
Краткая характеристика распространения.
Урогенитальные хламидиозы — наиболее распространенные формы хламидиозов. В крупных странах (США, Россия) ежегодно регистрируют несколько миллионов случаев, в мире — от 50 до 90 млн. случаев. Скудность начальных проявлений и тяжелые последствия урогенитальных хламидиозов, особенно у женщих (нарушения репродуктиной сферы, инфекционные осложнения), предъявляют особые требования к своевременной лабораторной диагностике. Хламидии сероваров D — K Chlamidia trachomatis, вызывающие урогенитальные хламидиозы, передаются от человека к человеку половым путем. Бессимптомное носительство наблюдается не менее чем у 5 % мужчин и 10% женщин. Отдельные серотипы этого возбудителя вызывают такие распространенные в прошлом заболевания как трахома (сопровождается поражениями конъюнктивы и прилегающих тканей глаза, часто приводит к катарактам и слепоте) ивенерическая лимфогранулема (регистрируют преимущественно в слаборазвитых странах Азии, Африки и Латинской Америки с теплым климатом).
Орнитоз — хламидийная инфекция, вызываемая C.psittaci. Человек заражается от птиц — основных хозяев этого возбудителя воздушно — пылевым и воздушно — капельным путем. В условиях города основную опасность представляют голуби (от 20% до 100% инфицировано этим возбудителем, чаще с ними контактируют дети. В домашних условиях источником могут быть канарейки и особенно попугаи (вызывают наиболее тяжелую форму — пситтакоз). Орнитоз часто протекает как тяжелая интерстициальная пневмония. Кроме этого, серотипы этого возбудителя вызывают зоонозные хламидиозы (например, так называемый вирусный аборт овец, хламидиозы крупного рогатого скота и др.), при контакте с больными животными могут развиваться различные формы хламидиозов у людей. Клинико — эпидемиологические особенности зоонозных хламидиозов у людей изучены недостаточно.
Бронхопневмонии , вызываемые C.pneumoniae. Это антропонозные инфекции, передаваемые от человека к человеку, большая часть случаев протекает субклинически. Возникают поражения верхних дыхательных путнй с последующим развитием бронхопневмонии. Это распространенные инфекции (антитела к C.pneumoniae выявляют почти у половины взрослого населения), однако диагностируются до настоящего времени плохо. С этим возбудителем связывают развитие отдельных форм бронхиальной астмы и атеросклероза.
С учетом многообразия клинических проявлений и необходимостью дифференциации различных клинических форм хламидиозов (прежде всего генитальных и экстрагенитальных) особое значение приобретает лабораторная диагностика.
XVIII. Хламидии
Хламидии – микроорганизмы со строгим внутриклеточным паразитизмом. Они впервые были обнаружены С. Провачеком и Л. Гальберштедтером в 1907 г. в отделяемом конъюнктивы при трахоме. Обнаруженные Провачеком микроколонии (тельца Гальберштедтера – Провачека) были окутаны мантией («хламидой»). С этого периода все микроорганизмы с аналогичными фенотипическими признаками стали относить к семейству Chlamydiaceae, роду Chlamydia.
Начиная с 1970 г. благодаря развитию ДНК-систематики, при изучении генома хламидий были использованы методы генетической рестрикции и молекулярной гибридизации, что позволило изучить структуру генов 16S и 23S рРНК у различных представителей порядка Chlamydiales. Результаты этих исследований (процент гомологии в последовательности генов 16S и 23S рибосомальной РНК), а также различия культуральных, биохимических, антигенных свойств хламидий, клинические проявления заболеваний и строго определенный круг хозяев явились предпосылкой к изменению номенклатуры и таксономии микроорганизмов, входивших в порядок Chlamydiales (К. Еwerett, 1999).
Классификация
В настоящее время выделено 4 семейства: Chlamydiacea, включающее 2 рода, патогенных для человека микроорганизмов – Chlamydia и Chlamydophila и 3 семейства родственных хламидиям микроорганизмов – Parachlamydiaceae, Simcaniacae и Waddliaceae.
Наибольшую роль в патологии человека из рода Chlamydia играет C. trachomatis, имееющая 15 сероваров. Серовары L-1, L-2, L-3 являются возбудителями пахового лимфогранулематоза; сероварианты А, Ва, В и С – возбудители трахомы. Остальные (от D до К) вызывают различные урогенитальные заболевания, пневмонию новорожденных, путь передачи – контактный.
Из рода Chlamydophila патогенными для людей являются C.pneumoniae, C. psittaci, C.abortus. C. pneumoniae имеет 4 сероварианта – TWAR, AR, KA, CWZ. Они вызывают пневмонии, ОРВИ, изучается их участие в патогенезе атеросклероза, бронхиальной астмы, саркоидоза. Путь передачи данных инфекций – воздушно-капельный.
C.abortus распространены среди животных, они колонизируют плаценту, вызывают аборты у животных, реже у женщин, работающих с овцами.
C. psittaci имеют восемь серовариантов. Они вызывают орнитоз (пситтакоз). Путь передачи – воздушно-капельный. Источник инфекции – птицы (для орнитоза – голуби, для пситтакоза – попугаи).
Морфология
Структура клеточной стенки хламидии соответствует общему принципу построения грамотрицательных бактерий. Она состоит из внутренней цитоплазматической и наружной мембран (обе являются двойными, обеспечивая прочность клеточной стенки). Антигенные свойства хламидий определяются внутренней мембраной, которая представлена липополисахаридами. В нее интегрированы белки наружной мембраны (Outer membrane proteins – ОМР). На основной белок наружной мембраны - Major Outer Membrane Protein (MOMP) приходится 60% общего количества белка. Остальная антигеннная структура представлена белками наружной мембраны второго типа – ОМР-2.
Все хламидии имеют общий групповой, родоспецифичный антиген (липополисахаридный комплекс, реактивной половиной которого является 2-кето-З-дезоксиоктановая кислота), используемый при диагностике заболевания иммунофлюоресцентными методами с применением специфических антител.
Белки MOMP и ОМР-2 содержат видо- и серотипоспецифические эпитопы. Однако в них имеются также области с высоким сходством среди видов (родоспецифические эпитопы), что обусловливает возможность появления перекрестных реакций. Основной белок клеточной мембраны и богатые цистеином другие белки связаны дисульфидными связями. Обнаружено пять генов дисульфидсвязанных изомераз, возможно играющих роль в реструктуризации цистеинбогатых белков при дифференциации элементарных телец в ретикулярные. У C. trachomatis выявлено 9 генов, кодирующих поверхностные мембранные белки, у C. pneumonia –18.
Хламидии и хламидофилы – строгие внутриклеточные патогены. Они способны синтезировать АТФ в незначительных количествах путем гликолиза и расщепления гликогена. Гликолитический цикл редуцирован, поскольку отсутствуют некоторые ферменты. Он компенсируется через пентозофосфатный и гексозофосфатные шунты. Хламидии в процессе приспособления к внутриклеточному паразитизму выработали уникальные структуры и биосинтетические механизмы, не имеющие аналогов у других бактерий. У хламидий не обнаружен высококонсервативный ген Ftsz, абсолютно необходимый для клеточного деления всех прокариот, поскольку он ответственен за образование клеточной перегородки во время деления клетки. У хламидий отсутствует пептидогликан – компонент клеточной стенки существующий как у грамположительных, так и у грамотрицательных бактерий, но при этом в геноме содержатся гены, кодирующие белки, которые необходимы для его полного синтеза.
Методом сканирующей электронной микроскопии на поверхности хламидий были выявлены куполообразные структуры, пронизанные микрофиламентами. Микрофиламенты выходят из центра, достигают мембраны включений и пронизывают ее. Функцию этой структуры связывают с транспортом питательных веществ от эукариотической клетки к паразиту. в геноме хламидий обнаружены гены, кодирующие аппарат для 3-го типа секреции, который обусловливает вирулентность грамотрицательных бактерий. Это образование осуществляет передачу сигнала от паразита к эукариотической клетке. В процесс взаимодействия хламидий с клеткой хозяина вовлечены не только поверхностные структуры хламидий, но и мембраны включений, поскольку в ассоциации с ними обнаружены хламидийные белки. У них отсутствуют цитохромы, т.е. они являются энергетическими паразитами. Культивируются в культурах клеток и желточном мешке куриного эмбриона.
Имеют уникальный двухфазный цикл развития: до проникновения в клетку хламидии инертны, это спороподобное состояние (элементарные тельца). Они обладают инфекциозностью, т.е. могут заражать клетки. Вначале элементарные тельца адгезируются на клетках-мишенях, а затем захватываются везикулами фагоцитов. В клетке везикулы с хламидиями двигаются в перинуклеарном пространстве и формируют включения внутри цитоплазмы. Элементарные тельца превращаются в большие метаболически активные неинфекционные ретикулярные тельца. Они оттесняют ядро клетки-хозяина к периферии. Сами располагаются около ядра и окружаются оболочкой клеточного происхождения «хламидой». Затем ретикулярное тельце делится, образуются дочерние формы, которые превращаются опять в элементарные тельца и выделяются из клетки.
Весь цикл развития занимает около трех суток. Процесс размножения хламидий является асинхронным и даже на клеточной стадии развития состоит из элементарных телец, ретикулярных телец и переходных форм. В инфицированной клетке не происходит слияние фагосомы с лизосомами, поэтому фагоцитоз носит незавершенный характер.
По химическому составу хламидии сходны с грамотрицательными бактериями, имеют в своем составе ДНК и РНК. Способны персистировать, могут переходить в L-формы, неподвижны, нет спор, у ретикулярных телец обнаружена микрокапсула.
Антигенная структура
Белковый термолабильный, находится в наружной мембране, проявляет свойства поринов. Он включает детерминанты, которые распознаются видо-, типо- и сероварспецифическими антителами; родоспецифический антиген (ЛПС) термостабилен и состоит из специфической детерминанты и групповой, дающей реакцию с некоторыми сальмонеллами.
Факторы патогенности
Имеют эндотоксин и экзотоксин, термолабильные мембранные эффекторы, которые связаны с типоспецифическими хламидийными антигенами и обеспечивают адгезию на клетках-мишенях, тропизм к определенным тканям. Выделяют нейраминидазу, которая разрушает сиаловую кислоту рецепторов, реагирующих с возбудителем. В результате увеличивается проницаемость тканей. Факторами патогенности являются и метаболиты, которые угнетают функции клеток хозяина.
Резистентность
Характеристика заболевания
Патогенез
Зависит от вида, сероварианта возбудителя, входных ворот и действия факторов патогенности. Ведущую роль в патогенезе хламидийной инфекции играют иммунопатологические механизмы.
Учитывая способность хламидий ингибироватъ слияние фагосом с лизосомами, фагоцитоз при хламидийной инфекции непродуктивный. При этом рост хламидий в моноцитах приостанавливается в промежуточном состоянии на стадии между элементарными и ретикулярными тельцами. На этом этапе в цитоплазме моноцитов обнаруживается липополисахарид клеточной стенки и отсутствует главный белок наружной мембраны (МОМР). Макрофаги презентирует Т-хелперам липополисахаридный (ЛПС) антиген и не презснтируют основной протективный антиген хламидий (МОМР). Иммунный ответ заведомо формируется к вариабельному ЛПС и оказывается неспецифическим по отношению к Chlamydia trachomatis.
Из одного общего предшественника Т-хелперов развиваются две функционально различающиеся, но взаимно регулируемые субпопуляции Т-клеток CD4+, продуцирующие собственные цитокины (табл. ).
Таблица