Клетки костной ткани
Костные клетки
Родоначальные клетки костной и хрящевой тканей
Костные клетки имеют мезенхимальное (мезенхимное, мезодермальное) происхождение. Во взрослом организме они образуются из остеогенных стволовых клеток-предшественников, которые локализуются на границе между костью и хрящевой или костномозговой тканью. Дифференцируясь, они превращаются в остеобласты, а затем - остеоциты. Рост длинных трубчатых костей осуществляется путем энхондрального окостенения. Причем увеличение диафизов в ширину происходит только со стороны периоста, а метафизов - только со стороны эндооста. Процесс костной резорбции имеет, соответственно, обратное направление (Burne, 1971, 1976; Фриденштейн, Лалыкина, 1973).
Схема образования костной и хрящевой ткани, построенная на основании работ А.Я. Фриденштейна, Е.А. Лурия (1980), А.Я. Фриденштейна и др. (1999), И.Л. Черткова, О.А. Гуревич (1984), В.П. Шахова (1996). Н. Castro-Malaspina et al., (1980, 1982) с некоторыми модификациями, представлена на рисунке.
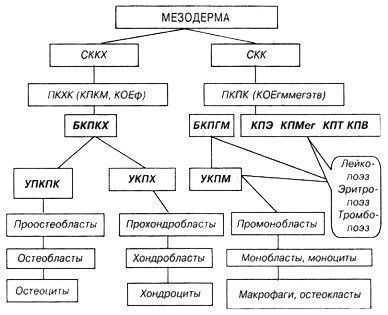
Схема остеогенеза, хондрогенеза и остеокластогенеза. СККХ - стволовая клетка костной и хрящевой ткани, СКК - стволовая клетка кроветворения, ПКПК - полипотентная клетка-предшественница кроветворной ткани, ПКХК - полипотентная клетка-предшественница для костной и хрящевой тканей, Б(У)КПКХ - би(уни)потентная клетка-предшественница костной и хрящевой ткани, КПКМ - клетка, переносящая кроветворное микроокружение, КОЕф - колониеобразующая единица фибробластов, У (Б) КПК (X, М, Г, Э, Мег, Т, В) - унипотентная (бипотентная) клетка-предшественница костной (хрящевой, макрфагальной, гранулоцитарной, эритроидной, мегакариоцитарной, Т и В-лимфоидной) ткани
Процесс образования костной ткани представляет собой сложный многоступенчатый процесс, при котором клетки различных гистогенетических линий проходят последовательную трансформацию путем пролиферации, дифференцировки и специализации с образованием композитной структуры, называемой костью.
При этом следует подчеркнуть, что если костная и хрящевая ткань формируется в эмбриогенезе из дорсального сомита мезодермы, то кроветворная ткань, из которой ведут свое происхождение остеокласты, - через стадию спланхнической мезодермы. По своему гистогенезу остеоциты и остеобласты ближе к соединительно-тканным, мышечным и кожным элементам, а остеокласты - к клеткам крови и эндотелию (Coalson, 1987). Наличие в остеокластобластомах эпителиальной и мышечной ткани, по-видимому, подтверждает эту точку зрения.
После расхождения направления развития остеохондрогенеза от гемопоэза в эмбриональном развитии, в зрелом организме процесс образования костных клеток осуществляется из более дифференцированного, фиксированного в тканях или циркулирующего незрелого стромального элемента (мезодермальной клетки, недифференцированного фибробласта, остеогенного предшественника или прекурсора) (Фриденштейн, Лурия, 1980; Альберст и др., 1994; Омельянченко и др., 1997). Наряду с наличием полипотентной стволовой клетки для костной и хрящевой ткани, существуют и более дифференцированные прекурсоры. СККХ имеют высокий пролиферативный потенциал, обладают полипотентностью. Они образуют, как минимум, костные и (или) хрящевые кариоциты, которые преимущественно находятся в G1-G2-стадии клеточного цикла (Фриденштейн, Лалыкина, 1977; Фриденштейн, Лурия, 1980; Фриденштейн и др., 1999; Чертков, Гуревич, 1984).
В культуре ткани in vivo и in vitro они образуют хрящевую или костную ткань, которая может быть представлена в форме колоний, обозначенных как колониеобразующие единицы фибробластов-КОЕф (Фриденштейн, Лурия, 1980). С помощью хромосомных и биохимических маркеров на радиационных химерах было показано, что КОЕф имеют клональную природу, отличную по своему происхождению от гемопоэтических клеток костного мозга, включая остеобласты и остеоциты (Чертков, Гуревич, 1984).
Нами в суспензионной культуре ткани костного мозга мышей линии Balb/c была изучена зависимость между количеством вводимых в среду кариоцитов и числом образовавшихся колоний. Для этого костный мозг вымывался в силиконизированную пробирку, суспендировался в D-MEM среде, содержащей 20% эмбриональной телячьей сыворотки, 40 мкг/мл гентамицина, 200 мМ, L-глютамина хепес и культивировался в течение 2-3 недель в пластиковых флаконах при 37 °С. Плотность посева составила от 104 до 107 клеток на мл.
Зависимость образования КОЕф при введении в культуру различного количества клеток костного мозга мышей линии Balb/c
| Количество вводимых клеток | Число образовавшихся колоний |
| 104 | 0 |
| 105 | 12 |
| 106 | 27 |
| 107 | 31 |
Приведенные данные свидетельствуют о том, что в целом зависимость между количеством вводимых в культуру миелокариоцитов и КОЕф носит линейный характер, что еще раз подтверждает их клональное происхождение.
При трансплантации их под капсулу почки или под кожу, они обладают способностью формировать костную, либо хрящевую ткань.
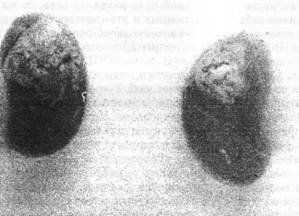
Макроскопический препарат эктопической костной ткани, выросшей под капсулой почки после трансплантации в нее костного мозга стрессированных мышей F1(CBAxC57Bl). Слева - на верхнем полюсе органа отчетливо виден большой очаг костеобразования. Справа - контроль (костный мозг взят от нестрессированного животного)
Одним из свойств СККХ является то, что они сохраняют свои пролиферативные и дифференцировочные потенции при многократном переносе первоначальной культуры от одного донора к другому. По-видимому, повреждение генома на этом уровне приводит к образованию остеосарком.
В результате дифференцировки СККХ образуются более дифференцированные клетки-предшественники типа КПКХ (клетки предшественники для костной и хрящевой ткани) или БКПКХ (бипотентные), затем - УПКПК и УПКПХ (унипотентные для кости, либо хряща). Общей закономерностью для пула родоначальных клеток любой ткани, в том числе и костной, является постепенное снижение способности к самообновлению и пролиферации, утрате полипотентности, увеличению доли прекурсоров, находящихся в S-периоде клеточного цикла, повышению чувствительности к действию ростовых факторов, гормонов, цитокинов и других регуляторных молекул. Теоретически этот процесс может идти равномерно или скачкообразно. Из-за этого течение остеогенеза может идти в различных режимах, темпе, с образованием костной ткани качественно и количественно отличной по своим морфофункциональным свойствам. На наш взгляд, введение биоматериала в кость обязательно включит тот или иной путь развития остеогенных клеток. Однако работ, выполненных в этом чрезвычайно интересном направлении, мы, к сожалению, не обнаружили.
Если ПКПКХ обладают полипотентностью, то БКПКХ образуют хрящевую или костную ткань, УКПК - только кость, а УКПХ - хрящ. Следует отметить, что все категории родоначальных клеток представляют собой чрезвычайно гетерогенную популяцию, внутри которой морфофункциональные свойства варьируют в широком диапазоне. Кроме того, для каждой из стадий развития КП имеется значительное количество переходных форм, которые все еще не могут быть идентифицированы с помощью имеющихся технологий. Несмотря на то, что методы выявления стромальных и остеогенных клеток-предшественников были открыты еще в начале 70-х годов, явного прогресса в понимании их свойств, способов регуляции и роли в процессах ремоделирования костной ткани достигнуто не было (Фриденштейн, Лалыкина, 1973; Фриденштейн и др., 1999; Чертков, Гуревич, 1984; Стецулла, Девятов, 1987; Омельянченко и др., 1997).
Следует отметить, что стволовые и коммитированные клетки-предшественники костной и хрящевой тканей находятся под контролем локальных и дистантных регуляторных механизмов. В последнюю группу входят факторы, которые оказывают свое действие через нейроэндокринную, иммунную, ретикулоэндотелиальную, опиатную, NO и другие системы путем выработки или связывания дальноранговых мессенжеров (эстрогены, глюкокортикоиды, эндорфины, адреналин и т.п.). Локальные механизмы действуют через прямое изменение морфофункциональных свойств микроокружения костной ткани, межклеточные контакты, местную выработку цитокинов, медиаторов, коротко-живущих биоактивных веществ и т.п. Межклеточные взаимодействия относятся к морфогенетическим процессам, они контролируют дифференцировку, специализацию, морфообразование клеток в тканях и органах. Механизмы их реализации осуществляются с помощью позиционно-информационных и индукционных взаимодействиях. Они еще малоизученны. Тем не менее, согласно концепции о позиционной информации, в организме существует морфогенетическое поле. Оно контролируется с помощью экспрессии гомейозисных генов типа НOХ1, НOХ2, НОХЗ, НOХ4, НOХ7, заставляя клетки помнить не только место своей локализации, в соответствии с координатными осями, но и выполнять миссию, которую они должны осуществить в процессе своей жизни, например восстановление кости при ее повреждении. Считается, что в сохранении позиционной информации большую роль играют мезенхимальные элементы, в частности макрофаги, остеобласты, остеоциты, остеокласты, эндотелий и фибробласты (Gilbert, 1994).
Индукционные механизмы регулируют процессы пролиферации и дифференцировки самообновляющихся клеточных популяций с помощью цитокинов, ростовых факторов, различных метаболитов и короткоранговых мессенжеров, вплоть до прямых клеточных взаимодействий.
Особенностью выбора направления дифференцировки поли- и бипотентных остеогенных предшественников является то, что он в первую очередь зависит от парциального давления кислорода. Если это давление достаточно высоко, то костные прекурсоры развиваются в направлении остеогенеза, а если низкое, то напротив, образуют хрящевую ткань (Bassett, Herman, 1961). При этом следует помнить, что адекватное поступление кислорода к клеткам возможно только при наличии развитой сети микроциркуляторного русла: максимальная величина удаления костных прекурсоров не должна превышать 100 мкм (Хэм, Кормак, 1983).
Система остеонов
Гаверсова система во взрослой кости постоянно обновляется. При этом всегда можно выделить несколько типов остеонов - эволюционирующих или развивающихся (5-10%), зрелых (50-75%), дегенерирующих или инволюционирующихся (10-20%), реконструирующихся (5-10%) и нежизнеспособных (5-10%).
Считается, что остеон (Гаверсова система) возникает только на основе туннеля, образующегося в результате действия моноцитов, макрофагов и остеокластов, заполняющегося изнутри концентрированными слоями костной ткани, формирующейся остеобластами и остеокластами (Хэм, Кормак, 1983). Следует отметить, что система остеонов представляет собой подвижную структуру, которая постоянно эволюционирует. Как это не парадоксально, работ, посвященных изучению кинетики остеонов, крайне мало. С помощью радионуклидных методов исследования было установлено, что годичная скорость замещения поверхностного слоя костной ткани составляет 5-10% (Harris, Heaney, 1969). По-видимому, и темп обновления остеонов имеет сходные параметры. Интересно, что диаметр остеонов в процессе развития не является постоянной величиной, а в течение всей своей жизни подвержен ряду последовательных изменений. Анализ литературных и собственных данных позволяет считать, что границы Гаверсовой системы, ограниченные линией цементации, у молодых, развивающихся и реконструирующихся остеонов составляют 80-150 мкм, зрелых - 120-300, а инволюцинирующих, дегенерирующих - менее 200 мкм. Если процесс образования остеонов протекает на границе надкостница/ кость, то вместо канала в начале формируется желобок, стенки которого выстланы остеогенными клетками, которые пролиферируют, формируя валик. Стенки этих клеточных выступов смыкаются, образуя полость, внутри которой, как правило, располагается не менее одной питающей артерии. Затем остеогенные клетки дифференцируются в остеобласты и остеоциты с формированием остеона. Предположения о том, что материал, используемый в травматологии, должен иметь диаметр пор равный размеру остеонов, высказывалось и ранее (Гюнтер и др., 1992). Однако этими авторами не был обоснован главный критерий, согласно которому размер пор должен соответствовать диаметру развивающихся, реконструирующихся, зрелых остеонов. При нарушении этого принципа в сторону увеличения или уменьшения диаметра пор полноценная костная ткань образовываться не будет. Иными словами, можно считать, что размер остеонов представляет собой важный морфообразующий фактор, который необходимо учитывать при создании искусственной костной ткани. Механизм этого феномена не совсем понятен. Он, вероятно, генетически запрограммирован в самих остеогенных клетках и является важным элементом костного микроокружения. Вместе с тем, следует подчеркнуть, что наряду с объемными характеристиками, например диаметром остеонов, при создании материалов необходимо учитывать и другие биологические принципы, речь о которых пойдет ниже.
А.В. Карпов, В.П. Шахов
Системы внешней фиксации и регуляторные механизмы оптимальной биомеханикиСтроение и виды костной ткани
19 Марта в 13:10 58907
Костная ткань представляет собой весьма совершенную специализированную разновидность тканей внутренней среды. В этой системе гармонично сочетаются такие противоположные свойства, как механическая прочность и функциональная пластичность, процессы новообразования и разрушения. Костная ткань состоит из клеток и межклеточного вещества, которые характеризуются определенной гистоархитектоникой. Основные клетки костной ткани — это остеобласты, остеоциты и остеокласты.Остеобласты имеют овальную или кубическую форму. Крупное светлое ядро располагается не в центре, оно несколько смещено к периферии цитоплазмы. Часто в ядре обнаруживается несколько ядрышек, что свидетельствует о высокой синтетической активности клетки.
Электронно-микроскопические исследования показали, что значительная часть цитоплазмы остеобласта заполнена многочисленными рибосомами и полисомами, канальцами гранулярной эндоплазматической сети, комплексом Гольджи, митохондриями, а также особыми матриксными пузырьками. Остеобласты обладают пролиферативной активностью, являются продуцентами межклеточного вещества и играют основную роль в минерализации костного матрикса. Они синтезируют и секретируют такие химические соединения, как щелочная фосфатаза, коллагены, остеонектин, остеопонтин, остеокальцин, костные морфогенетические белки и др. В матриксных пузырьках остеобластов содержатся многочисленные ферменты, которые, выделяясь за пределы клетки, инициируют процессы минерализации кости. Синтезируемый остеобластами органический матрикс костной ткани состоит преимущественно (90-95 %) из коллагена I типа, коллагенов III—V и других типов, а также из неколлагеновых белков (остеокальцин, остеопонтин, остеонектин, фосфопротеины, костные морфогенетические белки) и гликозаминогликановых субстанций. Белки неколлагеновой природы обладают свойствами регуляторов минерализации, остеоиндуктивных веществ, митогенных факторов, регуляторов скорости образования коллагеновых фибрилл. Тромбоспондин способствует адгезии остеобластов к поднадкостничному остеоиду кости человека. Остеокальцин считается потенциальным индикатором функции этих клеток. Ультраструктура остеобластов свидетельствует о том, что их функциональная активность различна. Наряду с функционально активными остеобластами, обладающими высокой синтетической активностью, имеются неактивные клетки. Чаще всего они локализуются на периферии кости со стороны костномозгового канала и входят в состав надкостницы. Строение таких клеток отличается малым содержанием органелл в цитоплазме.Остеоциты являются более дифференцированными клетками, чем остеобласты. Они имеют отростчатую форму. Отростки остеоцитов располагаются в канальцах, пронизывающих минерализованный костный матрикс в различных направлениях. Уплощенные тела остеоцитов находятся в специальных полостях — лакунах — и со всех сторон окружены минерализованным костным матриксом. Значительную часть цитоплазмы остеоцита занимает овоидное ядро. Органеллы синтеза в цитоплазме развиты слабо: имеются немногочисленные полисомы, короткие канальцы эндоплазматической сети, единичные митохондрии. В связи с тем что канальцы соседних лакун анастомозируют друг с другом, отростки остеоцитов связаны между собой при помощи специализированных щелевых контактов. В небольшом пространстве вокруг тел и отростков остеоцитов циркулирует тканевая жидкость, содержащая определенную концентрацию Са2+ и РО43-, могут содержаться неминерализованные или частично минерализованные коллагеновые фибриллы.
Функция остеоцитов заключается в сохранении целостности костного матрикса за счет участия в регуляции минерализации костной ткани и обеспечения ответа на механические стимулы. В настоящее время накапливается все больше данных о том, что эти клетки принимают активное участие в метаболических процессах, протекающих в межклеточном веществе кости, в поддержании постоянства ионного баланса в организме. Функциональная активность остеоцитов в значительной мере зависит от стадии их жизненного цикла и действия гормональных и цитокиновых факторов.Остеокласты — это крупные многоядерные клетки с резко оксифильной цитоплазмой. Они являются частью фагоцитарно-макрофагальной системы организма, производными моноцитов крови. На периферии клетки определяется гофрированная щеточная каемка. В цитоплазме обнаруживается много рибосом и полисом, митохондрий, канальцев эндоплазматической сети, хорошо развит комплекс Гольджи. Отличительной особенностью ультраструктуры остеокластов является наличие большого количества лизосом, фагосом, вакуолей и везикул.
Остеокласты обладают способностью создавать локально у своей поверхности кислую среду в результате интенсивно идущих в этих клетках процессов гликолиза. Кислая среда в области непосредственного контакта цитоплазмы остеокластов и межклеточного вещества способствует растворению минеральных солей и создает оптимальные условия для действия протеолитических и ряда других ферментов лизосом. Цитохимическим маркером остеокластов служит активность изофермента кислой фосфатазы, который называется кислой нитрофенилфосфатазой. Функции остеокластов заключаются в резорбции (разрушении) костной ткани и участии в процессе ремодуляции костных структур в ходе эмбрионального и постнатального развития. Межклеточное вещество костных тканей состоит из органического и неорганического компонентов. Органические соединения представлены коллагенами I, III, IV, V, IX, XIII типов (около 95 %), неколлагеновыми белками (костные морфогенетические белки, остеокальцин, остеопонтин, тромбоспондин, костный сиалопротеин и др.), гликозаминогликанами и протеогликанами. Неорганическая часть костного матрикса представлена кристаллами гидроксиапатита, содержащими в большом количестве ионы кальция и фосфора; в значительно меньшем количестве в его состав входят соли магния, калия, фториды, бикарбонаты. Межклеточное вещество кости постоянно обновляется. Разрушение старого межклеточного вещества представляет собой достаточно сложный и еще не ясный во многих деталях процесс, в котором принимают участие все типы клеток костной ткани и ряд гуморальных факторов, но особенно заметную и важную роль играют остеокласты.Типы костной ткани. В зависимости от микроскопического строения различают две основные разновидности костной ткани — ретикулофиброзную (грубоволокнистую) и пластинчатую.
Ретикулофиброзная костная ткань широко представлена в эмбриогенезе и раннем постнатальном гистогенезе костей скелета, а у взрослых встречается в местах прикрепления сухожилий к костям, по линии зарастания черепных швов, а также в области переломов. Как в эмбриогенезе, так и при регенерации ретикулофиброзная костная ткань с течением времени всегда замещается пластинчатой. Характерным в строении ретикулофиброзной костной ткани является неупорядоченное, диффузное расположение костных клеток в межклеточном веществе. Мощные пучки коллагеновых волокон слабо минерализованы и идут в различных направлениях. Плотность расположения остеоцитов в ретикулофиброзной костной ткани более высокая, чем в пластинчатой, и они не имеют определенной ориентации по отношению к коллагеновым (оссеиновым) волокнам.
Пластинчатая костная ткань является основной тканью в составе практически всех костей человека. В этой разновидности костной ткани минерализованное межклеточное вещество образует особые костные пластинки толщиной 5-7 мкм. Каждая костная пластинка представляет собой совокупность близко расположенных друг к другу параллельных коллагеновых волокон, пропитанных кристаллами гидроксиапатита. В соседних пластинках волокна располагаются под разными углами, что придает кости дополнительную прочность. Между костными пластинками в лакунах упорядоченно лежат костные клетки — остеоциты. Отростки остеоцитов по костным канальцам проникают в окружающие их пластинки, вступая в межклеточные контакты с другими костными клетками. Различают три системы костных пластинок: окружающие (генеральные, бывают наружными и внутренними), концентрические (входят в структуру остеона), вставочные (представляют собой остатки разрушающихся остеонов).
В составе кости различают компактное и губчатое вещество. Оба они образованы пластинчатой костной тканью. Особенности гистоархитектоники пластинчатой кости будут представлены далее при описании кости как органа. Болезни суставовВ.И. Мазуров
Клетки костной ткани описание
В состав скелета любого взрослого человека входит 206 различных костей, все они различны по строению и роли. На первый взгляд они кажутся твердыми, негибкими и безжизненными. Но это ошибочное впечатление, в них непрерывно происходят различные обменные процессы, разрушение и регенерация. Они, в совокупности с мышцами и связками, образуют особую систему, что носит название «костно-мышечная ткань», основная функция которой — опорно-двигательная. Она образована из нескольких видов особых клеток, которые различаются по структуре, функциональным особенностям и значению. О костных клетках, их строение и функциях далее и пойдет речь.
Строение костной ткани
Это отдельный вид соединительной ткани, из нее образуются все кости в человеческом теле. В ее состав входят особые клетки и межклеточное вещество. Последнее включает органический матрикс, состоящий из коллагеновых волокон (90-95% от общей массы) и минеральных компонентов, в основном солей кальция (5-10%). Благодаря такому составу костная ткань человека имеет гармоничное сочетание твердости и эластичности. Различают три группы клеток: остеокласты (слева), остеобласты (посередине), остеоциты (справа на фото).
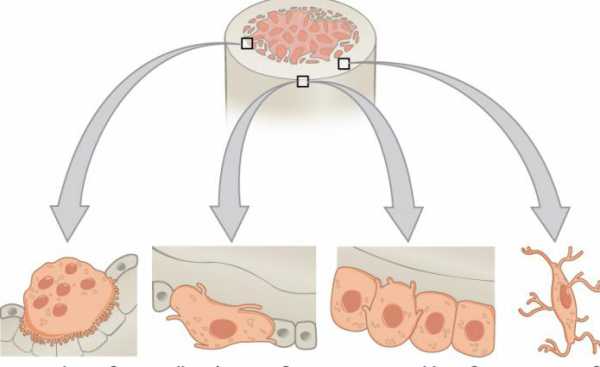
Более подробно остановимся на них далее. Коллаген, содержащийся в матриксе, имеет отличия от своих аналогов, находящихся в других тканях, главным образом за счет того, что содержит больше специфических полипептидов. Волокна расположены, как правило, параллельно уровню наиболее вероятных нагрузок на кость. Именно благодаря нему сохраняется эластичность и упругость.
Если кость подвергнуть действию соляной кислоты, то минеральные вещества будут растворены, а вот органические (оссеин) останутся. Они сохранят форму, но станут чрезмерно гибкими и сильно подверженными деформированию. Такое состояние характерно для маленьких детей. У них высоко содержание оссеина, поэтому кости более эластичны, чем у взрослых. И обратный случай, когда теряются органические вещества, но остаются минеральные. Это происходит, если, к примеру, кость обжечь: она сохранит свою форму, но приобретет вместе с тем сильную хрупкость и может разрушиться даже от незначительного прикосновения. Такие изменения состав костной ткани претерпевает в старости. Доля минеральных солей доходит до 80% от всей массы. Поэтому пожилые люди более подвержены различного рода переломам и травмам.
Если установить плотность костной ткани (объем), то это позволит оценить прочность скелета и его отдельных частей. Такие исследования проводятся с использованием компьютерной томографии. Своевременная диагностика позволяет начать лечение или поддерживающую терапию вовремя.
Остеобласты (активные): особенности строения
Остеобласты – это клетки костной ткани, располагающиеся в верхних ее слоях, имеющие многоугольную, кубическую форму с различного вида отростками. Внутреннее содержимое мало чем отличается от других. Хорошо развитый зернистый эндоплазматический ретикуллум содержит различные элементы, рибосомы, аппарат Гольджи, округлой или овальной формы ядро богатое хроматином и содержащее ядрышко. Снаружи эти клетки костной ткани окружены тончайшими микрофибриллами.
Главная функция остеобластов – синтез компонентов межклеточного вещества. Это коллаген (преимущественно первого типа), гликопротеины матрикса (остеокальцин, остеонектин, остеопонтин, костный сиалопротеин), протеогликаны (бигликан, гиалуроновая кислота, декорин), а также различные костные морфогенетические белки, факторы роста, ферменты, фосфопротеины. Нарушение выработки всех этих соединений остеобластами наблюдается при некоторых заболеваниях. Например, недостаток витамина С (цинга) у детей характеризуется нарушением развития и роста костей вследствие дефекта синтеза коллагена и гликозаминогликанов. По этой же причине и замедляется восстановление костной ткани, заживление при переломах. Так как остеобласты фактически отвечают за рост, то присутствуют исключительно в развивающейся костной ткани.
Механизм минерализации остеобластами органического матрикса
Существует два способа:
- Отложение кристаллов гидроксилата вдоль фибрилл коллагена из перенасыщенной внеклеточной жидкости. Особую роль при этом отводят некоторым протеогликанам, которые связывают кальций и удерживают его в зонах зазоров.
- Секреция особых матричных пузырьков. Это мелкие мембранные структуры, которые синтезируются и выделяются остеобластами. В них в большой концентрации содержится фосфат кальция и щелочная фосфатаза. Особая микросреда, создаваемая внутри пузырьков, благоприятствует образованию первых гидроксиапатитовых кристаллов.
Скорость минерализации остеоида (костная ткань на стадии формирования) может существенно меняться, в норме она занимает около 15 суток. Нарушения могут происходить при снижении концентрации ионов кальция в крови или фосфата. Результатом этого является размягчение и деформация костей – остеомаляция. Аналогичные нарушения наблюдаются, например, при рахите (дефицит витамина D).
Неактивные (покоящиеся) остеобласты
Они образуются из активных остеобластов, у нерастущей кости покрывают около 80-95% ее поверхности. Они имеют уплощенную форму с веретеновидным ядром. Остальные органеллы редуцированы. Но сохраняются рецепторы, реагирующие на различные гормоны и факторы роста. Между покоящимися остеобластами и остеоцитами сохраняется связь и таким образом образуется система, регулирующая минеральный обмен. Если происходит какое-либо повреждение (травмы, переломы), то они активизируются, и начинается активный синтез коллагена, выработка органического матрикса. Другими словами, за счет их происходит регенерация костных тканей. В то же время они могут быть причиной злокачественной опухоли – остеосаркомы.
Остеоциты: строение и функции
Эти клетки составляют основу зрелой костной ткани. Форма у них веретенообразная, с множеством отростков. Органелл значительно меньше по сравнению с остеобластами, есть округлое ядро (в нем преобладает гетеохроматин) с ядрышком. Остеоциты располагаются в лакунах, но непосредственно с матриксом не соприкасаются, а окружены тонким слоем костной жидкости. За счет нее осуществляется питание клеток.
Аналогично отделены и их отростки, имеющие достаточно большую длину до 50 мкм, располагающиеся в специальных канальцах. Их очень много, костная ткань буквально пронизана ими, они образуют ее дренажную систему, в которой и содержится тканевая жидкость. Через нее осуществляется обмен веществ между межклеточным веществом и клетками. Также стоит отметить, что они не делятся, а образуются из остеобластов и являются основными компонентами в сформировавшейся костной ткани.
Основная функция остеоцитов – поддержание нормального состояния костного матрикса и баланса кальция и фосфора в организме. Они способны воспринимать механические напряжения, и чувствительны к электрическим потенциалам, возникающим при действии деформирующих сил. Реагируя на них, они запускают локальный процесс, при котором соединительная костная ткань начинает перестраиваться.
Остеокласты
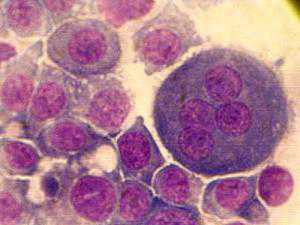
Такое название получили крупные клетки, содержащие от 5 до 100 ядер, имеющие моноцитарное происхождение, разрушающие кости и хрящи или, по-другому, вызывающие их резорбцию. В цитоплазме остеокластов содержится много митохондрий, элементов ЭПС (зернистой) и аппарат Гольджи, рибосомы, а также различные по функции лизосомы. В ядрах содержится большое количество хроматина и есть хорошо различимые ядрышки. Также имеется достаточное количество цитоплазматических отростков, больше всего их располагается на поверхности, прилегающей к разрушаемой кости. Они увеличивают площадь соприкосновения с ней. Костная ткань начинает разрушаться при повышении уровня особого гормона (паратиреоидного), который приводит к активации остеокластов. Механизм этого процесса связывают с выделением ими углекислого газа, который под воздействием специального фермента (карбоангидраза) превращается в кислоту, имеющую название угольная, она и растворяет соли кальция.
Механизм резорбции костной ткани
Стоит отметить, что процесс разрушения протекает циклически, и периоды высокой активности каждой клетки неизменно сменяются периодами покоя. Резорбция протекает в несколько этапов:
- Прикрепление остеокласта к разрушаемой поверхности кости, при этом наблюдается выраженная перестройка его цитоскелета.
- Окисление содержимого лакун. Это происходит либо путем выделения в них содержимого вакуолей, имеющего кислую среду, либо в результате действия протонных насосов.
- Разрушение минерального компонента матрикса.
- Растворение органических соединений в результате действия ферментов, секретируемых остеокластами в лакуну и активированными кислой средой.
- Выведение продуктов разрушения костной ткани.
Регуляция деятельности остеокластов определяется общими и местными факторами. К первым, например, относятся паратгормон, витамин D, они стимулируют активность. А угнетающими являются кальцитонин и эстрогены. К местным относится такой фактор, как создание электрического локального поля при механическом напряжении, к которому эти клетки очень чувствительны.
Строение грубоволокнистой костной ткани
Второе ее название — ретикулофиброзная. Она формируется у зародыша, как будущая основа костей. У взрослого же человека ее присутствие минимально, она сохраняется в швах черепа после того, как они зарастают и в зонах, где сухожилия прикрепляются к костям, а также в участках остеогенеза, например, при заживлении различного рода переломов. Строение костной ткани этого вида специфическое. Коллагеновые волокна собраны в плотные пучки, которые расположены неупорядоченно, имеют между собой «перекладины». Она обладает низкой механической прочностью, содержание остеоцитов значительно выше по сравнению с пластинчатой разновидностью. В патологических условиях наращивание костной ткани этого типа происходит при переломе кости или при болезни Педжета.
Особенности пластинчатой костной ткани
Она образована костными пластинками, имеющими толщину 4-15 мкм. Они, в свою очередь, состоят их трех компонентов: остеоцитов, основного вещества и коллагеновых тонких волокон. Из этой ткани образованы все кости взрослого человека. Волокна коллагена первого типа лежат параллельно относительно друг друга и ориентированы в определенном направлении, у соседних же костных пластинок они направлены в противоположную сторону и перекрещиваются практически под прямым углом. Между ними находятся тела остеоцитов в лакунах. Такое строение костной ткани обеспечивает ей наибольшую прочность.
Губчатое вещество кости
Встречается также название «трабекулярное вещество». Если проводить аналогию, то структура сравнима с обычной губкой, построенной из костных пластинок с ячейками между ними. Расположены они упорядоченно, в соответствии с распределенной функциональной нагрузкой. Из губчатого вещества в основном построены эпифизы длинных костей, часть смешанных и плоских и все короткие. Видно, что в основном это легкие и в то же время прочные части скелета человека, которые испытывают нагрузку в различных направлениях. Функции костной ткани находятся в прямой взаимосвязи с ее строением, которое в данном случае обеспечивает большую площадь для метаболических процессов, осуществляемых на ней, придает высокую прочность в совокупности с небольшой массой.
Плотное (компактное) вещество кости: что это?
Из компактного вещества состоят диафизы трубчатых костей, кроме того, оно тонкой пластинкой покрывает их эпифизы снаружи. Его пронизывают узкие каналы, через них проходят нервные волокна и кровеносные сосуды. Некоторые из них располагаются параллельно костной поверхности (центральные или гаверсовы). Другие выходят на поверхность кости (питательные отверстия), через них внутрь проникают артерии и нервы, а наружу — вены. Центральный канал, в совокупности с окружающими его костными пластинками, образует так называемую гаверсову систему (остеон). Это основное содержимое компактного вещества и их рассматривают как его морфофункциональную единицу.
Остеон – структурная единица костной ткани
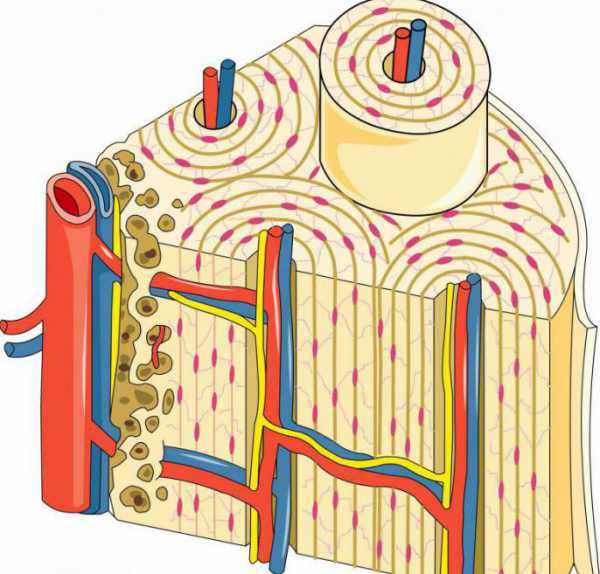
Второе его название — гаверсова система. Это совокупность костных пластинок, имеющих вид цилиндров вставленных друг в друга, пространство между ними заполняют остеоциты. В центре располагается гаверсов канал, через него проходят обеспечивающие обмен веществ в костных клетках кровеносные сосуды. Между соседними структурными единицами есть вставочные (интерстициальные) пластинки. По сути, они являются остатками остеонов, существовавших ранее и разрушившихся в тот момент, когда костная ткань претерпевала перестройку. Также существуют еще генеральные и окружающие пластинки, они образуют самый внутренний и наружный слой компактного вещества кости соответственно.
Надкостница: строение и значение
Исходя из названия, можно определить, что она покрывает кости снаружи. Прикрепляется она к ним с помощью коллагеновых волокон, собранных в толстые пучки, которые проникают и сплетаются с наружным слоем костных пластинок. Имеет два выраженных слоя:
- наружный (его образует плотная волокнистая, неоформленная соединительная ткань, в ней преобладают волокна, располагающиеся параллельно к поверхности кости);
- внутренний слой хорошо выражен у детей и менее заметен у взрослых (образован рыхлой волокнистой соединительной тканью, в которой есть веретенообразные плоские клетки – неактивные остеобласты и их предшественники).
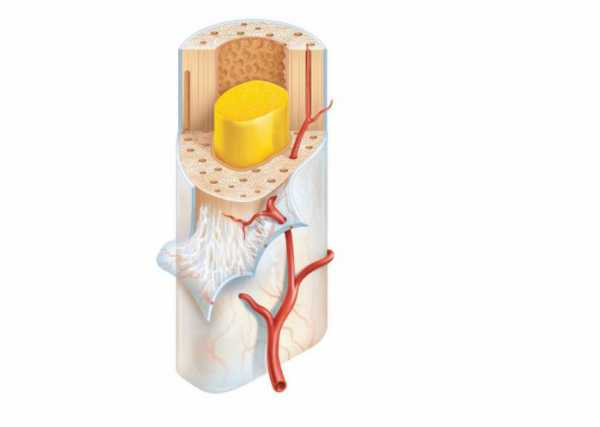
Надкостница выполняет несколько важных функций. Во-первых, трофическую, то есть обеспечивает кость питанием, поскольку на поверхности содержит сосуды, которые проникают внутрь вместе с нервами через специальные питательные отверстия. Эти каналы питают костный мозг. Во-вторых, регенераторную. Она объясняется наличием остеогенных клеток, которые при стимуляции трансформируются в активные остеобласты, вырабатывающие матрикс и вызывающие наращивание костной ткани, обеспечивающие ее регенерацию. В-третьих, механическую или опорную функцию. То есть обеспечение механической связи кости с другими прикрепляющимися к ней структурами (сухожилиями, мышцами и связками).
Функции костной ткани
Среди основных функций можно перечислить следующие:
- Двигательная, опорная (биомеханическая).
- Защитная. Кости оберегают от повреждений головной мозг, сосуды и нервы, внутренние органы и т. д.
- Кроветворная: в костном мозге происходит гемо — и лимфопоэз.
- Метаболическая функция (участие в обмене веществ).
- Репараторная и регенераторная, заключающиеся в восстановлении и регенерации костной ткани.
- Морфобразующая роль.
- Костная ткань – это своеобразное депо минеральных веществ и ростовых факторов.
Иммунология и биохимия
Кость не инертная ткань, а динамически обновляющаяся в течение всей жизни соединительная ткань. Старая матрица кости постоянно заменяется новообразованной матрицей. Этот непрерывный процесс называется ремоделированием кости. Ремоделирование имеет большое значение для поддержания объема костной ткани и её прочности. Объем костной ткани поддерживается за счет баланса резорбции костной ткани и формированием костной ткани. Костные клетки состоят из клеток линии остеобластов и линии остеокластов. Их дифференцировка и функции регулируются остеотропными гормонами и цитокинами. Недавние исследования показали, что клетки линии остеобластов участвуют не только в формировании костной ткани, но и в резорбции кости, поддерживая дифференциацию и активацию остеокластов. В этой статье отражены данные морфологических характеристик клеток линии остеобластов, их функции и дифференцировка. Остеобласты
Остеобласты - это кубовидные клетки, расположенные вдоль поверхности кости, составляют 4-6% от общего числа резидентных костных клеток, основная функция - формирование кости. Морфологические характеристики остеобластов отражают их способность к синтезу белка: обильная шероховатая эндоплазматической сеть, видимый аппарат Гольджи, различные секреторные везикулы. Как поляризованные клетки, остеобласты секретируют остеоид матрицы кости.
Остеобласты образуются из мезенхимальных стволовых клеток (МСК). Коммитирование МСК в клетки предшественники остеогенеза требует экспрессии специфических генов, синтез костных морфогенетических белков и членов специфического сигнального пути.
Синтез костной матрицы остеобластами включает два основных этапа: осаждение органической матрицы и ее последующая минерализация. На первом этапе остеобласты секретируют коллаген, в основном типа I коллаген, неколлагеновые белки (остеонектин, сиалопротеин II и остеопонтин) и протеогликаны, включая декорин и бигликан, которые образуют органическую матрицу. После этого в два этапа происходит минерализация костной матрицы: везикулярная и фибриллярная фазы. В везикулярную фазу из апикальной части остеобластов освобождаются матричные пузырьки (везикулы, диаметр 30 - 200 нм), содержащие ионы кальция, связанные с протеогликанами и другими органическими компонентами, имеющими отрицательный заряд. Когда остеобласты секретируют ферменты, которые разрушают протеогликаны, ионы кальция высвобождаются из протеогликанов в кальциевые каналы матрицы мембраны везикул. Эти каналы образованы белками, называемыми аннексины.
С другой стороны, фосфатсодержащие соединения пузырьков гидролизуются щелочной фосфатазой, секретируемой остеобластами, высвобождая ионы фосфата внутри матричных везикул. Ионы фосфата и кальция внутри везикул связываются, образуя кристаллы гидроксиапатита. Фибриллярная фаза возникает тогда, когда перенасыщение ионами кальция и фосфата в матричных пузырьках приводит к разрыву этих структур, и кристаллы гидроксиапатита распространяются в окружающую матрицу.
Зрелые остеобласты могут подвергаться
- апоптозу, стать
- остеоцитами или
- костными обкладочными клетками.
Обкладочные клетки кости в линии остеобластов
Костные обкладочные клетки в состоянии покоя представляют собой плоские остеобласты, которые покрывают поверхность кости в местах, где отсутствуют процессы синтеза и резорбции костной ткани. Эти клетки обладают тонким и плоским ядерным профилем; в цитоплазме содержится несколько органелл - эндоплазматический ретикулум и комплекс Гольджи.
Секреторная активность костных обкладочных клеток зависит от физиологического состояния костной ткани. Эти клетки могут приобретать секреторную активность, увеличивая размер, и принимать внешний вид шестигранника. Функции костных подкладочных клеток не полностью изучены, но было показано, что эти клетки при резорбции кости предотвращают непосредственное взаимодействие остеокластов с костным матриксом, участвуют дифференцировке остеокластов, освобождая остеопротегерин и активатор рецепторов ядерного фактора (RANKL). Кроме того, обкладочные клетки кости вместе с другими костными клетками являются важным компонентом анатомической структуры цикла ремоделирования костной ткани.
Остеоциты в линии остеобластов
Остеоциты составляют 90-95% от общего количества костных клеток, являются наиболее распространенными и долгоживущими клетками, с продолжительностью жизни до 25 лет. В течение многих десятилетий из-за трудностей в выделении остеоцитов из костной матрицы их ошибочно относили к пассивным клеткам. Развитие новых технологий позволило установить, что эти клетки играют многочисленные важные функции в кости. Остеоциты расположены в лакунах, окруженных минерализованной костной матрицей, имеют морфологию дендритных клеток. Морфология остеоцитов зависит от типа кости. Например, остеоциты губчатой кости более округлые, чем остеоциты кортикальной кости удлиненной морфологии.Остеоциты образуются при дифференцировке остеобластов. В этом процессе выделяют четыре узнаваемых этапа: остеоид-остеоцитарный, преостеоцитарный, молодой остеоцитарный и зрелый остеоцитарный. В конце цикла формирования кости субпопуляции остеобластов становятся остеоцитами, включенными в матрицу кости. Этот процесс сопровождается заметными морфологическими и ультраструктурными изменениями, включая уменьшение круглого размера остеобластов. Количество органелл, таких как шероховатая эндоплазматическая сеть и комплекс Гольджи, уменьшается, а отношение ядро/цитоплазма возрастает, снижается синтез белка и секреция. Зрелые остеоциты полностью погружены внутри минерализованной костной матрицы. Исчезают маркеры остеобластов, появляются маркеры остеоцитов, включая дентин матричного белка 1 и склеростин. В то время как тело остеоцита находится внутри лакуны, его цитоплазматические дендриты (до 50 на каждую клетку) пересекают крошечные туннели, которые берут начало из пространства лакун, под названием канальцы, образуя остеоцит - лакуноканаликулярную систему. Остеоциты системы обмениваются информацией, как с помощью щелевых контактов, так и с помощью малых сигнальных молекул, таких как простагландины и оксид азота. Кроме того, остеоцит - лакуноканаликулярная система находится в непосредственной близости от сосудов, в результате чего кислород и питательные вещества достигают остеоцитов. Межклеточная связь также достигается за счет межклеточной жидкости, которая течет между отростками остеоцитов и канальцев. Остеоцит-лакуноканаликулярная система остеоцитов действуют как механосенсоры, обладает способностью обнаруживать механические нагрузки и тем самым способствовать адаптации кости к ежедневным механическим силам. Этим образом, остеоциты, вероятно, действуют как дирежеры ремоделирования кости, посредством регулирования активности остеобластов и остеокластов. Кроме того, апоптоз остеоцитов рассматривается как хемотаксический сигнал к резорбции костной ткани остеокластами. Показано, что во время резорбции кости апоптотические остеоциты поглощаются остеокластами. Механочувствительная функция остеоцитов достигается за счет стратегического расположения этих клеток в костной матрице. В остеоцитах механические стимулы переводятся в биохимические сигналы. Это явление называется пьезоэлектрический эффект. Механизмы и компоненты, с помощью которых остеоциты преобразуют механические стимулы в биохимические сигналы, не очень хорошо известны. При механической стимуляции остеоциты производят несколько вторичных мессенджеров, таких как АТФ, оксид азота (NO), Са2+, и простагландины (PGE2 и PGI2), которые влияют на физиологию кости. Вне зависимости от вовлеченного механизма, важно отметить, что механочувствительная функция остеоцитов возможна благодаря сложной канальцевой сети, которая обеспечивает связь между костными клетками.